Эмбриология. Дробление
Эмбриональный период имеет следующие этапы: дробление, гаструляция, гисто- и органогенез.
. Дробление. В результате дробления образуется бластула;
. Гаструляция. В результате гаструляции образуются зародышевые листки;
. Гисто- и органогенез - образование тканей и осевых органов зародыша.
При личиночной форме онтогенеза эмбриональный период начинается с образования зиготы и заканчивается выходом из яй-
цевых оболочек. При неличиночной форме онтогенеза этот период начинается с образования зиготы и заканчивается выходом из зародышевых оболочек. При внутриутробной форме онтогенеза - с образования зиготы до рождения особи.
Дробление
В результате оплодотворения образуется зигота, которая начинает дробиться. Дробление сопровождается митотическими делениями. Между делениями в короткой интерфазе отсутствует пост- митотический период, а синтез ДНК начинается в телофазе предыдущего митотического деления. Не происходит роста зародыша. Суммарный объем зародыша не изменяется. Клетки, образующиеся в процессе дробления, называются бластомерами, а зародыш - бластулой. Тип дробления зависит от количества и распределения желтка в яйцеклетке (рис. 67).
Рис. 67. Типы яиц и способы их дробления (яйца и зародыши изображены вверх анимальным полюсом).
I - изолецитальное яйцо (а) и его равномерное дробление (б); II и III - телолецитальные яйца (в, д), и их дробления - дискоидальное (г) и неравномерное (е); IV - центролецитальное яйцо (ж) и его поверхностное дробление (з).
Типы дробления
Дробление может быть полным равномерным, полным неравномерным, неполным дискоидальным, поверхностным (рис. 68).
Полное равномерное дробление характерно для изолецитальных яиц - например для ланцетника. Ядро зиготы делится митозом на

Рис. 68. Типы яиц и соответствующие им типы дробления.
два, затем делится цитоплазма. Борозда дробления проходит по меридиану, образуются два бластомера. Затем снова делится ядро и на поверхности зародыша появляется вторая борозда дробления, идущая по меридиану, перпендикулярно первой. Образуются 4 бластомера, 3-я борозда проходит по экватору и делит его на 8 частей. Затем происходит чередование меридионального и экваториального дроблений. Число бластомеров увеличивается. Зародыш на стадии 32 бластомеров называется морулой. Дробление продолжается до образования зародыша, похожего на пузырек, стенки которого образованы одним слоем клеток, называемых бластодермой. Бластомеры расходятся от центра зародыша, образуя полость, которая называется первичной, или бластоцелью. Бластомеры имеют одинаковый размер. В результате такого дробления образуется целобластула.
Полное неравномерное дробление свойственно телолецитальным яйцам с умеренным содержанием желтка, например у лягушек. 1-я и 2-я борозды дробления проходят по меридианам и полностью делят яйцо на 4 части, 3-я борозда смещена в сторону анимального полюса, где нет желтка. Бластомеры имеют неодинаковую величину: на анимальном полюсе они меньше (микромеры), на вегетативном больше (макромеры). Желток затрудняет дробление, и поэтому дробление макромеров идет медленнее, чем микромеров. Стенка бластулы состоит из нескольких рядов клеток. Первичная полость мала и смещена к анимальному полюсу. Образуется амфибластула.
Неполному дискоидальному дроблению подвергаются телолецитальные яйца с большим содержанием желтка, например у рептилий, птиц. Дробление идет только на анимальном полюсе. 1-я и 2-я борозды дробления проходят по меридиану перпендикулярно друг другу, 3-я борозда смещена к анимальному полюсу. В результате этого образуется зародышевый диск. Бластоцель располагается под слоем бластодермы в виде щели. Бластула называется дискобластулой.
Неполное поверхностное дробление характерно для центролецитальных яиц, например для членистоногих. Ядра центролециталь-
ных яиц многократно делятся и перемещаются к периферии, где в цитоплазме нет желтка. Цитоплазма образует бластомеры. Бласту- ла имеет один слой бластомеров. Бластоцель заполнена желтком. Такая бластула называется перибластулой.
Гаструляция
По окончании периода дробления у многоклеточных животных наступает период образования зародышевых листков - гаст- руляция. Гаструляция связана с перемещением эмбрионального материала. Сначала образуется ранняя гаструла, имеющая два зародышевых листка (эктодерму и энтодерму) , затем поздняя гаструла, когда формируется 3-й зародышевый листок - мезодерма. Зародыш, образовавшийся в результате гаструляции, называется гаструлой.
Образование ранней гаструлы происходит следующими способами:
. иммиграцией (выселением клеток), у кишечнополостных;
. инвагинацией (впячиванием), у ланцетника;
. эпиболией (обрастанием), у лягушки;
. деляминацией (расщеплением), у кишечнополостных.
При иммиграции часть клеток бластодермы с поверхности зародыша уходит в бластоцель. Образуется наружный слой - эктодерма и внутренний - энтодерма. Бластоцель заполнена клетками.
При инвагинации определенный участок бластодермы (вегетативный полюс) прогибается внутрь и достигает анимального полюса. Образуется двуслойный зародыш-гаструла. Наружный слой клеток - эктодерма, внутренний - энтодерма выстилает полость первичной кишки (гастроцель). Отверстие, при помощи которого полость сообщается с внешней средой, называется первичным ртом - бластопором. У первичноротых животных (черви, моллюски, членистоногие) он превращается в ротовое отверстие, у вторичноро- тых - в анальное отверстие, а рот образуется на противоположном конце (хордовые).
Эпиболия характерна для животных, которые развиваются из телолецитальных яиц. Образование гаструлы идет за счет быст-
рого размножения микромеров, которые обрастают вегетативный полюс. Макромеры оказываются внутри зародыша. Образование бластопора не происходит и нет гастроцели.
Деляминация встречается у кишечнополостных, бластула которых похожа на морулу. Клетки бластодермы делятся на наружный и внутренний слои. Наружный слой образует эктодерму, внутренний - энтодерму (рис. 69).
У всех многоклеточных, кроме губок и кишечнополостных, возникает 3-й зародышевый листок - мезодерма. Формирование мезодермы происходит двумя способами: телобластическим или энтероцельным.
Телобластический способ свойствен первичноротым. На границе между эктодермой и энтодермой по бокам от бластопора

Рис. 69. Типы гаструл.
а - инвагинационная гаструла; б, в - две стадии развития иммиграционной гаструлы; г, д - две стадии развития деляминационной гаструлы; е, ж - две стадии развития эпиболической гаструлы; 1 - эктодерма; 2 - энтодерма; 3 - бластоцель.
клетки - телобласты - начинают делиться и дают начало мезодерме.
Энтероцельный способ характерен для вторичноротых. Клетки, формирующие мезодерму, обособляются в виде карманов первичной кишки. Полости карманов превращаются в целом. Мезодерма делится на отдельные участки - сомиты, из которых образуются определенные ткани и органы.
Гисто- и органогенез
После образования мезодермы начинается процесс гисто- и органогенеза. Сначала формируются осевые органы - нервная трубка, хорда, затем все остальные органы (рис. 70).
У ланцетника из эктодермы на спинной стороне зародыша образуется нервная трубка. Остальная эктодерма формирует кожный эпителий и его производные. Из энто- и мезодермы под нервной трубкой формируется хорда. Под хордой находится кишечная трубка, по бокам от хорды - мезодерма сомитов. Наружная часть сомита, прилегающая к эктодерме, называется дермотомом. Из нее формируется соединительная ткань кожи. Внутренняя часть - склеротом - дает начало скелету. Между дермотомом и склеротомом находится миотом, дающий начало поперечно-полосатой мускулатуре. Под сомитами находятся ножки (нефрогонотом), из которых формируется мочеполовая система.
Целомические мешки формируются симметрично по бокам. Стенки целомических мешков, обращенные в сторону кишечника, на-

Рис. 70. Образование мезодермы (по Шимкевичу, 1925, модифицировано).
а - у первичноротых; б - у вторичноротых;
1 - эктодерма; 2 - мезенхима; 3 - энтодерма; 4 - телобласт (а) и целомическая мезодерма (б).
зывают спланхноплеврой, в сторону эктодермы - соматоплеврой. Эти листки участвуют в образовании сердечно-сосудистой системы, плевры, брюшины, перикарда (рис. 71).
Таким образом, из эктодермы формируются кожный эпителий, кожные железы, эмаль зубов, волосы, ногти, когти.
Из энтодермы образуются эпителий средней кишки, печень, поджелудочная железа, щитовидная железа, тимус, эпителий дыхательной системы.
Мезодерма участвует в образовании мышц, соединительной ткани, костной ткани, каналов выделительной системы, кровеносной системы, части ткани половых желез.
На основе ряда существенных характеристик (полнота, равномерность и симметрия деления) выделяют ряд типов дробления . Типы дробления во многом определяются распределением веществ (в том числе, желтка ) по цитоплазме яйца и характером межклеточных контактов, которые устанавливаются между бластомерами.
Дробление может быть
полным (голобластическим) или неполным (меробластическим),
равномерным (бластомеры более-менее одинаковы по величине) и
неравномерным (бластомеры не одинаковы по величине, выделяются две - три размерные группы, обычно называемые макро- и микромерами), наконец по характеру симметрии различают радиальное, спиральное, различные варианты билатеризованных и анархическое дробление. В каждом из этих типов выделяют ряд вариантов.
По степени полноты деления:
Голобластическое дробление
Плоскости дробления разделяют яйцо полностью. Выделяют полное равномерное дробление, при котором бластомеры не различаются по размерам (такой тип дробления характерен для гомолецитальных яиц), и полное неравномерное дробление, при котором бластомеры могут существенно различаться по размерам. Такой тип дробления характерен для умеренно телолецитальных яиц.
Меробластическое дробление Дискоидальное
ограничено относительно небольшим участком у анимального полюса,
плоскости дробления не проходят через всё яйцо и не захватывают желток.
Такой тип дробления типичен для телолецитальных яиц богатых желтком (птицы , рептилии ). Такое дробление называют также дискоидальным , так как в результате дробления на анимальном полюсе образуется небольшой диск клеток (бластодиск ).
Дискоидальное дробление (от греч. dískos - диск и éidos - вид), один из типов дробления яиц у животных с телолецитальными яйцами (скорпионы, головоногие моллюски, хрящевые и костистые рыбы, пресмыкающиеся и птицы). При Дискоидальном дробление делится лишь небольшой диск относительно свободной от желтка и содержащей ядро цитоплазмы.
Поверхностное
ядро зиготы делится в центральном островке цитоплазмы ,
получающиеся клетки перемещаются на поверхность яйца , образуя поверхностный слой клеток (бластодерму ) вокруг лежащего в центре желтка.
Такой тип дробления наблюдается у членистоногих .
По типу симметрии дробящегося яйца
Радиальное
Ось яйца является осью радиальной симметрии. Типично для ланцетника , осетровых , амфибий , иглокожих , круглоротых.
Спиральное
В анафазе бластомеры разворачиваются. Отличается лево-правой дисимметрией (энантиоморфизм) уже на стадии четырёх (иногда двух) бластомеров. Типично для некоторых моллюсков , кольчатых и ресничных червей .
Билатеральное
Имеется 1 плоскость симметрии. Типично для аскариды .
Анархическое
Бластомеры слабо связаны между собой, сначала образуют цепочки. Типично для кишечнополостных .
Энуклеация - в гистологии удаление клеточного ядра.
Эпибласт = Эктодерма, иначе эктобласт, иначе эпибласт - наружный пласт зародыша Metazoa, а равно и наружный слой стенки тела низших Metazoa (многоклеточных).
Дидактическая единица №3 – Законы генетики
АЛЛОПОЛИПЛОИДИЯ (от алло... и полиплоидия ) - наследственное изменение в клетках растений, реже животных, заключающееся в кратном увеличении числа наборов хромосом при межвидовых или межродовых скрещиваниях. Встречается в природе и может быть получена целенаправленно (ржано-пшеничные, капустно-редечные гибриды). Имеет важное значение в процессах видообразования у растений.
Анемия Фанкони
(G. Fanconi, род. в 1892 г., швейц. педиатр ; син. Фанкони синдром ) наследственная болезнь , характеризующаяся гипоплазией костного мозга, панцитопенией, а также аномалиями развития кожи (гиперпигментация ), костной системы (недоразвитие 1 пястной или лучевой кости) и (или) внутренних органов (почек, селезенки); наследуется по аутосомно-рецессивному типу.
ДНК-гираза когда расплетает, она спираль сдвигает на границе репликона и образуется суперспираль, необычайно крутая раскрутка. Чтобы её ликвидировать появляются свевилазы , которые способны произвести надрезание в местах сверхскрутки в районе фосфатного мостика. Формируется шарнир Кернса и через него суперспираль сбрасывается. Появляется фермент хеликаза (SSB), который стабилизирует её, сам биосинтез начинается с синтеза праймера (затравки), причем затравка состоит из РНК. Появляется особый мультиферментный комплекс - праймосома. В ней 3 фермента: 1). Праймаза (синтезирует РНК-затравку) 2). ДНК-белок (ДНК зависимая рибонуклеоизид 3 фосфатаза) 3). N"-белок (ДНК зависимая отефаза) ДНК-белок и N"-белок определяет начало репликации, с которой и начинается затравка. Обычно это кусочек из 6 нуклеотидов. Далее праймосома перемещается на соседнюю точку Ори. Первая нить 3"5" - лидирующая, а вторая 5"3" - запаздывающая. После этого к праймеру присоединяется ДНК-полимераза-3. Начинается рост цепи ДНК - элонгация, которая идет до терминирующих кодонов, находящихся у точки Ори соседнего репликона. После этого появляется фермент РНКаза, которая удаляет праймер, при этом образуется пустота, её застраивает фермент ДНК-полимераза-1. После биосинтеза у соседних репликонов образуются фрагменты Оказаки , сливающиеся воедино за счет ДНК-лигазы. После этого в этом участке ДНК происходит восстановление спиральности. Это происходит за счет топоизомеразы-3, которая представляет смесь гиразы и w-белка. Таким образом происходит полный синтез ДНК. В итоге ДНК репликация идет следующим образом: 1). ДНК-гираза раскручивает участок ДНК-репликон, начиная с точки Ори. 2). На границе создается суперспираль. 3). ДНК- свевилаза создает шарнир Кернса и суперспираль сбрасывается. 4). Хеликаза (SSB) стабилизирует одноцепочечные нити не давая слиться. 5). Праймосома. Мобильный промотор репликации за счет проймазы ДНА-белка создает РНК затравку в точке Ори. 6). ДНК-полимераза -3 синтезирует молекулу ДНК - фрагмент Оказаки. 7). РНКаза удаляет праймер затравку. 8). ДНК-полимераза-1 застраивает Брешь 9). ДНК - Лигаза сшивает фрагменты Оказаки 10). Топоизомераза-3 формирует спираль.
Перетяжки, разделяющие дробящуюся яйцеклетку на все более мелкие клетки (бластомеры), называются бороздами дробления. Дробление это многократные митотические деления зиготы, в результате которых зародыш становится многоклеточным, не меняя при этом существенно своего объема. Образование многоклеточности первая и основная биологическая роль дробления. Вторая роль состоит в восстановлении ядерно-плазматического отношения, которое падает в ходе стадии большого роста ооцита. Видовые особенности процесса дробления определяются двумя основными параметрами: количеством и распределением желточных белков в цитоплазме (желток подавляет дробление); присутствием в цитоплазме факторов, которые влияют на ориентацию митотического веретена и время его образования. Дробление начинается вскоре после оплодотворения и заканчивается, когда у зародыша достигается новое равновесие между ядром и цитоплазмой. Дробление строго координированный процесс, находящийся под генетическим контролем.
В большинстве других случаев клеточной пролиферации в период между митозами происходит рост клеток. Клетка увеличивается в объеме почти вдвое и затем делится. Такой рост приводит к увеличению общего объема клеток при сохранении относительно постоянного отношения объема ядра к объему цитоплазмы. В период дробления зиготы объем цитоплазмы не возрастает: огромная масса цитоплазмы зиготы разделяется на все более мелкие клетки. Это деление цитоплазмы яйца, не сопровождающееся ростом, осуществляется путем выпадения G1-периода в интерфазе, тогда как митозы следуют друг за другом с большой скоростью.
Темпы увеличения числа клеток в период дробления намного выше, чем на стадии гаструляции. Одним из последствий высокой интенсивности делений в процессе дробления является постепенное уменьшение отношения объема цитоплазмы к объему ядра. Изменение скорости, с которой происходит снижение отношения объема цитоплазмы к объему ядра, у многих типов зародышей является решающим фактором, определяющим время активации некоторых генов. От митотического деления соматических клеток дробление отличается тем, что полученные в результате дробления клетки не растут, а поэтому с каждым следующим делением становятся всё более мелкими, при этом увеличивается только их количество, а зародыш в целом не растет. Получающиеся при дроблении клетки мало дифференцированы и сравнительно однородны.
Дробление (сегментация) у отдельных представителей разряда позвоночных имеет в общем одинаковое течение; однако, как уже было упомянуто выше, оно находилось под влиянием факторов, которые во время филогенеза воздействовали на развитие в виде последствий влияния внутренней и внешней среды, в которой организмы проживали во время своего родового развития (ценогенетические факторы).
При наблюдении за изменениями , происходящими в яйцеклетках согласно филогенетическому развитию яиц отдельных представителей разряда позвоночных, можно заметить, что яйцевые клетки в значительной мере отличаются друг от друга по содержанию питательного и строительного вещества - желтка. Яйцевые клетки ланцетника (Amfioxus), организма, который в филогенетическом отношении считается наиболее низко организованным существом, но который уже обладает прочной спинной областью, относятся к числу олиголецитальных.
Однако, в соответствии с филогенетическим развитием , количество желтка в яйцеклетках позвоночных животных, являющихся филогенетически наиболее высоко организованными организмами, все более возрастает, достигая максимального количества в птичьих яйцеклетках, которые являются относительно очень крупными и полилецитальными. Под влиянием ценогенетических факторов (факторов, воздействующих из внешней среды и обусловливаемых изменением образа жизни, а следовательно, и развития) количество желтка в процессе филогенетического развития по направлению к человеку все более уменьшается, благодаря чему яйцеклетки человека и высших млекопитающих становятся снова (вторично) олиголецитальными.
Наличие вариабельного количества желтка
оказывает, как уже было сказано выше, значительное влияние на процесс дробления яйцеклетки. Яйцевые клетки с малым содержанием желтка (олиголецитальные) дробятся полностью, то есть все вещество оплодотворенного яйца при дроблении делится на новые клетки, бластомеры (яйцеклетки голобластического вида). Наоборот, у яйцеклеток, содержащих желтка больше, или даже большое количество желтка (полилецитальных), борозды дробления непрерывно дробят только меньшую часть ооплазмы, расположенную на так называемом анимальном полюсе, где желточных гранул меньше (яйцеклетки меробластического вида).
В соответствии с этим у отдельных представителей разряда
позвоночных различаются следующие типы дробления.
1. Полное дробление . К полному, тотальному дроблению относятся те случаи, когда в процессе дробящего деления делится вся оплодотворенная яйцевая клетка и борозды дробления распространяются по всей ее поверхности. По этому типу дробятся яйцевые клетки голобластического вида. В зависимости от содержания в ооплазме большего или меньшего количества желтка, а также в зависимости от его распределения в ооплазме, при дроблении возникают бластомеры либо сравнительно одинаковой величины (полное равномерное, эквальное, или адэквальное дробление), либо бластомеры различной величины, а именно более крупные в области с большим содержанием желтка и менее крупные в том месте, где желтка меньше (полное неравномерное, инэквальное дробление). Более крупные бластомеры называются макромерами, менее крупные - микромерами.
Полное эквальное, или адэквальное , дробление свойственно олиголецитальным, изолецитальным яйцеклеткам (ланцетник, высшие млекопитающие и человек); по полному инэквальному типу дробятся мезолецитальные яйцевые клетки анизолецитального и умеренно телолецитального вида (некоторые низшие рыбы и земноводные).
2. Частичное, парциальное, дробление . По частичному типу дробятся яйцевые клетки, содержащие значительное количество желтка (полилецитальные яйцеклетки), у которых из-за их больших размеров борозды дробления при клеточном делении проникают только в область анимального полюса, где находится клеточное ядро и где слой ооплазмы содержит меньше желточных гранул (высшие рыбы, пресмыкающиеся, птицы и некоторые низшие млекопитающие, яйцеродные).
При таком дроблении на анимальном полюсе сравнительно крупного яйца дробится только круглое поле (диск), в то время как остаток яйцевой клетки (желточный шар) остается не раздробленным (парциальное дисковидное дробление). У насекомых их полилецитальные центролецитальные яйцеклетки хотя и дробятся по всей поверхности, но центр клетки, содержащий большое количество желтка, остается не раздробленным (парциальное поверхностное дробление).
Строение бластулы
Дробление приводит к образованию шарообразного зародыша – бластулы . Если образуется сплошной шар без полости внутри, то такой зародыш называют морулой. Образование бластулы или морулы зависит от свойств цитоплазмы. Бластула образуется при достаточной вязкости цитоплазмы, морула – при слабой вязкости. При достаточной вязкости цитоплазмы бластомеры сохраняют округлую форму и только в местах соприкосновения слегка сплющиваются. Вследствие этого между ними появляется щель, которая по мере дробления увеличивается, заполняется жидкостью и превращается в бластоцель. При слабой вязкости цитоплазмы бластомеры не округляются и располагаются тесно друг возле друга, щели нет и полость не образуется. Бластулы различны по своему строению и зависят от типа дробления.
Типы бластул
Различают пять типов бластул : целобластулу, амфибластулу, стерробластулу, дискобластулу и перибластулу. Целобластула образуется при полном равномерном дроблении из яйцеклеток гомолецитального типа (ланцетник). Бластодерма целобластулы состоит из одного ряда более или менее одинаковых бластомеров, внутри находится крупная полость – бластоцель.
Бластодерма амфибластулы состоит из нескольких рядов клеток. Бластодерма в анимальной части тоньше, чем в вегетативной. Бластоцель меньших размеров, чем у ланцетника, и смещена к анимальному полюсу. Такого типа бластула образуется при полном неравномерном дроблении и характерна для круглоротых и земноводных.
Стерробластула состоит из одного ряда крупных бластомеров, которые глубоко заходят в полость бластулы, бластоцель в связи с этим или очень малая, или отсутствует (некоторые членистоногие).
Дискобластула образуется при неполном дискоидальном дроблении. Бластоцель в виде узкой щели находится между зародышевым диском и желтком. Крыша бластулы представлена бластодермой, а дно желтком. Такая бластула характерна для костистых рыб, пресмыкающихся и птиц. Бластодерма перибластулы состоит из одного ряда клеток, которые окружают желток. Полость в ней отсутствует. Перибластула наблюдается у некоторых насекомых.
По мере такого дробления эм-бриональные клетки , называемые бластомерами, становятся мельче (палинтомическое деление). Этот процесс называется дроблением . Оно продолжается до формирова-ния мелкоклеточного зародыша.
Типы дробления оплодотворенных яиц обусловлены их строением. Дробление может быть полным и частичным (рис. 26), поверхностным , радиальным , спиральным и билатеральным .
При полном , или голобластическом , дроблении вся зигота или яй-цеклетка делится на бластомеры, и борозды дробления, проникают в самые глубокие части яйца. Иначе говоря, борозды дробления разде-ляют яйцо на бластомеры полностью. В зависимости от того, какой размер имеют получившиеся бластомеры, дробление может быть рав-номерным и неравномерным . При равномерном дроблении у гомолецитальных яиц длительное время сохраняется синхронность дробле-ния бластомеров, поэтому они имеют одинаковые размеры и наблюда-ется удвоение их количества: 2, 4, 8, 16, 32, 64. При неравномерном дроблении синхронность дробления нарушена, так как бедные желт-ком бластомеры дробятся быстрее бластомеров, богатых желтком. В результате они оказываются разных размеров и последовательного удвоения их количества не происходит.
При частичном , или меробластическом , дроблении борозды не разделяют яйцо полностью, поэтому большая его часть не дробится. Здесь также есть свои варианты. Например, у птиц и рептилий заро-дыш развивается за счет незначительного количества цитоплазмы, расположенной у анимального (верхнего) полюса. Остальная часть яйца заполнена желтком и не дробится. Образуется цитоплазматиче-ский диск, по которому и проходят борозды дробления.
Поверхностное дробление характерно для насекомых , имеющих центролецитальные яйца. При этом несколько первых делений ядра, находящегося в центре яйца, происходят без деления цитоплазмы. За-тем ядра окружаются небольшим слоем цитоплазмы и как бы всплы-вают из массы желтка на поверхность, объединяясь там с поверхност-ным слоем цитоплазмы. В этой зоне и идет образование бластомеров, внутри же остается неразделившийся желток.
В зависимости от расположения бластомеров относительно друг друга выделяют три типа дробления: радиальное , спиральное и билатеральное .
При радиальном дроблении два первых деления являются меридио-нальными, затем следуют экваториальные деления, когда плоскость деления перпендикулярна к плоскостям первых двух делений. В связи с этим четыре бластомера анимального полушария лежат над четырь-мя бластомерами вегетативного полушария, что создает радиальную симметрию (рис. 27, а).
Радиальное дробление яиц свойственно та-ким группам животных, как иглокожие. У морского ежа даже на стадии 64 бластомеров идентифицировать отдельные из них невозможно, так как первые этапы дробления дают одинаковые клетки.
При спиральном дроблении (рис. 27, б) анимальные клетки смеща-ются по отношению к анимально-вегетативной (передне-задней) оси яйца. Плоскость дробления проходит под углом к ней и к экватору яй-ца. Оси митотических веретен образуют спирали, и бластомеры как бы чередуются друг с другом. Спиральное дробление характеризуется жесткой упорядоченностью, что позволяет проследить дальнейшую судьбу бластомеров, вернее, их производных. После первых двух дроблений образуются четыре бластомера A , B , C и D . Последующие деления дают бластомеры меньшего размера (микромеры), 1 a , 1 b , 1 c и 1 d . Следующие микромеры обозначают 2 a , 2 b , 2 c и 2 d и т. д. Установ-лено, что у ряда животных, дробление яиц которых осуществляется по спиральному типу, вся эктодерма происходит от трех квартетов мик-ромеров, отделяющихся от первых четырех бластомеров в ходе после-довательных этапов дробления.
Многим группам беспозвоночных животных, таким, в частности, как немертины, кольчатые черви , моллюски , присуще спиральное дробление. Иногда высказывается мнение, что такой тип дробления является признаком филогенетического родства между ними. С уве-ренностью можно утверждать только то, что распространение спи-рального дробления в нескольких типах говорит лишь о том, что это — консервативный признак. Материал с сайта
Биологическое значение дробления
- Переход к многоклеточности
- Увеличение ядерно-цитоплазматического отношения
Характерные черты дробления
Дробление как особый этап онтогенеза животных имеет характерные черты, которые свойственны большинству животных, но могут отсутствовать у некоторых групп.
- Бластомеры делятся очень быстро (у дрозофилы - раз в 20 минут) и более или менее синхронно.
- Интерфаза сокращена до S-периода; в связи с этим транскрипция собственных генов зародыша полностью подавлена, транскрибируются только запасённые в яйцеклетке материнские мРНК.
- Между делениями нет периода роста, так что общая масса зародыша не растёт.
По всем этим характеристикам дробление млекопитающих резко отклоняется от типичного. Бластомеры делятся у них медленно, синхронность нарушается уже после 1-2 делений, в это же время активируется собственный геном зародыша.
Классификация типов дробления
На основе ряда существенных характеристик (степень детерминированности, полнота, равномерность и симметрия деления) выделяют ряд типов дробления . Типы дробления во многом определяются распределением веществ (в том числе, желтка) по цитоплазме яйца и характером межклеточных контактов, которые устанавливаются между бластомерами.
Дробление может быть: детерминированным и регулятивным; полным (голобластическим) или неполным (меробластическим); равномерным (бластомеры более-менее одинаковы по величине) и неравномерным (бластомеры не одинаковы по величине, выделяются две - три размерные группы, обычно называемые макро- и микромерами); наконец, по характеру симметрии различают радиальное, спиральное, различные варианты билатеризованных и анархическое дробление. В каждом из этих типов выделяют ряд вариантов.
По степени детерминированности
Детерминированное
Недетерминированное (регулятивное)
(Бластомеры тотипотентны)По степени полноты делений
Голобластическое дробление
Плоскости дробления разделяют яйцо полностью. Выделяют полное равномерное дробление, при котором бластомеры не различаются по размерам (такой тип дробления характерен для гомолецитальных и алецитальных яиц), и полное неравномерное дробление, при котором бластомеры могут существенно различаться по размерам. Такой тип дробления характерен для умеренно телолецитальных яиц.
Меробластическое дробление
- Дискоидальное
- ограничено относительно небольшим участком у анимального полюса,
- плоскости дробления не проходят через всё яйцо и не захватывают желток.
Такой тип дробления типичен для телолецитальных яиц, богатых желтком (птицы , рептилии). Такое дробление называют также дискоидальным , так как в результате дробления на анимальном полюсе образуется небольшой диск клеток (бластодиск).
- Поверхностное
- ядро зиготы делится в центральном островке цитоплазмы ,
- получающиеся ядра перемещаются на поверхность яйца , образуя поверхностный слой ядер (синцитиальную бластодерму) вокруг лежащего в центре желтка. Затем ядра разделяются мембранами, и бластодерма становится клеточной.
Такой тип дробления наблюдается у членистоногих .
По типу симметрии дробящегося яйца
Радиальное
Билатеральное
Имеется 1 плоскость симметрии. Типично для аскариды .
Анархическое
Бластомеры слабо связаны между собой, сначала образуют цепочки или бесформенную массу; часто у одного вида встречаются разные варианты расположения бластомеров. Типично для кишечнополостных .
Литература
- Белоусов Л.В. Основы общей эмбриологии. - Москва: Издательство Московского университета: Наука, 2005. - ISBN 5-211-04965-9
- Токин Б.П. Общая эмбриология: Учеб. для биол. спец. ун-тов. - 4-е изд., перераб. и доп. - М.: Высш. шк., 1987. - 480 с.
| Эмбриогенез | |
|---|---|
| Биология развития | |
| Стадии | Зигота Морула Бластула (Организмы: Диплобласты Триплобласты) Бластоциста Гаструла Нейрула Эмбрион |
| Процессы | Дробление яйца Бластуляция Гаструляция (деламинация , инвагинация , иммиграция , эпиболия) Нейруляция Органогенез |
| Зародышевые листки | Эктодерма Энтодерма Мезодерма |
| Дифференцировка клеток | Бластомер Эмбриобласт Трофобласт Эпибласт Гипобласт |
Wikimedia Foundation . 2010 .
Смотреть что такое "Дробление (эмбриология)" в других словарях:
Дробление: Дробление (технология) измельчение твёрдого тела до определенного размера; Дробление (полиграфия) воспроизведение на оттиске одного и того же печатающего элемента дважды, со смещением; Дробление (эмбриология) ряд… … Википедия
Дробление: Дробление (технология) измельчение твёрдого тела до определенного размера; Дробление (полиграфия) воспроизведение на оттиске одного и того же печатающего элемента дважды, со смещением; Дробление (эмбриология) ряд последовательных… … Википедия
- (от древнегреческого ἔμβρυον, зародыш, «эмбрион»; и λογία, логия) это наука, изучающая развитие зародыша. Зародышем называют любой организм на ранних стадиях развития до рождения или вылупления, или, в случае растений, до момента прорастания.… … Википедия
Дробление: эмбрион млекопитающего. z.p. zona striata, p.gl полярные тельца, a. двуклеточная стадия, b. четырёхклеточная стадия, c. восьмиклеточная стадия, d, e. морула Дробление ряд последовательных митотических делений оплодотворенного… … Википедия
дробление радиальное - ЭМБРИОЛОГИЯ ЖИВОТНЫХ ДРОБЛЕНИЕ РАДИАЛЬНОЕ – первое дробление меридиональное, проходит в меридиональной плоскости яйца. Второе дробление также меридиональное, оно проходит через главную ось яйца, но под прямым углом к плоскости первого дробления.… …
дробление анархическое - ЭМБРИОЛОГИЯ ЖИВОТНЫХ ДРОБЛЕНИЕ АНАРХИЧЕСКОЕ [БЕСПОРЯДОЧНОЕ, ХАОТИЧНОЕ] – дробление яиц метагенетических медуз – Oceania armata. Первая борозда дробления меридиональная, врезающаяся, появляется она на анимальном полюсе. Вторая борозда тоже… … Общая эмбриология: Терминологический словарь
дробление гетероквадратное - ЭМБРИОЛОГИЯ ЖИВОТНЫХ ДРОБЛЕНИЕ ГЕТЕРОКВАДРАТНОЕ – дробление аннелид, моллюсков, немертин, планарий. Неравномерное спиральное дробление, когда клетки основного квартета (первые четыре бластомера) неодинаковы по величине, то и их производные также… … Общая эмбриология: Терминологический словарь
дробление асинхронное - ЭМБРИОЛОГИЯ ЖИВОТНЫХ ДРОБЛЕНИЕ АСИНХРОННОЕ – дробление яиц телолецитальных (амфибия). Деление вегетативных бластомеров происходит медленней, по сравнению с бластомерами анимального полюса … Общая эмбриология: Терминологический словарь
дробление билатеральное - ЭМБРИОЛОГИЯ ЖИВОТНЫХ ДРОБЛЕНИЕ БИЛАТЕРАЛЬНОЕ [ДВУСТОРОННЕ СИММЕТРИЧНОЕ] – дробление яиц нематод, коловраток, асцидий. Характеризуется появлением билатеральной симметрии в расположении бластомеров уже на ранних этапах дробления. Каждый бластомер… … Общая эмбриология: Терминологический словарь
дробление голобластическое - ЭМБРИОЛОГИЯ ЖИВОТНЫХ ДРОБЛЕНИЕ ГОЛОБЛАСТИЧЕСКОЕ [ПОЛНОЕ] – дробление яиц алецитального (плоские черви), изолецитального типа (ланцетник) и некоторых яйцевых клеток телолецитального типа (амфибии). Дробятся все части зиготы. При дроблении вся… … Общая эмбриология: Терминологический словарь
 Прапорщики и другие воинские звания: как они появились на самом деле Младший прапорщик в ссср
Прапорщики и другие воинские звания: как они появились на самом деле Младший прапорщик в ссср Можгинское медицинское училище министерства здравоохранения удмуртской республики Можгинское медицинское училище поступление
Можгинское медицинское училище министерства здравоохранения удмуртской республики Можгинское медицинское училище поступление Правильные многоугольники презентация к уроку по геометрии (9 класс) на тему Все о правильный многоугольник презентация
Правильные многоугольники презентация к уроку по геометрии (9 класс) на тему Все о правильный многоугольник презентация Таблица персонажи эпохи дворцовых переворотов
Таблица персонажи эпохи дворцовых переворотов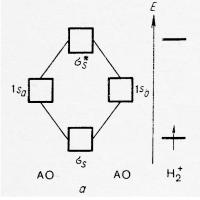 Метод молекулярных орбиталей (ММО)
Метод молекулярных орбиталей (ММО) Герундий - примеры использования - учим испанский язык
Герундий - примеры использования - учим испанский язык Методика определения ионов кальция и магния в природных водах
Методика определения ионов кальция и магния в природных водах